|
| source ici |
Le moins qu’on puisse dire, c’est que les découvertes de Waddington dans les années 1950 sur les liens étranges entre génétique et environnement n’ont pas vraiment eu d’écho à l'époque. Il faut dire que ça tombait pile au moment où l'on découvrait l’ADN et l’image fascinante de programme génétique orienta durablement la recherche en biologie vers des modèles réductionnistes de type un gène = une enzyme. Richard Lewontin rapporte ainsi (dans la triple hélice) que le biologiste moléculaire Sydney Brenner affirma un jour que “s’il disposait de la séquence d’ADN complète d’un organisme et d’un ordinateur suffisamment puissant, il saurait calculer l’organisme”!
Ce n’est qu’au milieu des années 1990 que l’on s’intéressa de nouveau aux expériences de Waddington. Susan Lindquist en particulier chercha à comprendre les raisons moléculaires pour lesquelles on observe autant de mutations chez des organismes (animaux ou végétaux) soumis à des stress pendant leur phase de croissance, et leur lien avec l'évolution des espèces. Une recherche tellement fructueuse qu'elle lui valut la médaille nationale de la science en 2010...
HSP90 usual suspect
Ce n’est qu’au milieu des années 1990 que l’on s’intéressa de nouveau aux expériences de Waddington. Susan Lindquist en particulier chercha à comprendre les raisons moléculaires pour lesquelles on observe autant de mutations chez des organismes (animaux ou végétaux) soumis à des stress pendant leur phase de croissance, et leur lien avec l'évolution des espèces. Une recherche tellement fructueuse qu'elle lui valut la médaille nationale de la science en 2010...
HSP90 usual suspect
Dans une expérience de 1998 devenue célèbre (ici en pdf), Lindquist démontra d'abord que si l'on inhibe chimiquement une protéine appelée HSP90 durant l'état larvaire de la drosophile, on observe chez l'adulte la même avalanche d’anomalies que lorsqu’on la soumet à un intense stress thermique ou alimentaire:
 |
| (Source: Rutherford & Lindquist 1998, pdf) Exactement comme dans l’expérience de Waddington, Lindquist découvrit que ces mutations sont en grande partie héréditaires, même lorsque les descendants de la lignée mutante retrouvent un fonctionnement normal de leurs protéines HSP90. Ah, ah! Tiendrait-on le coupable moléculaire? Des baby-sitter en forme de protéine (ou l’inverse) |
 |
| La molécule HSP90 (source CNRS) |
Avant de répondre, il me faut vous en dire un tout petit plus sur le fonctionnement des protéines et le rôle de cette fameuse HSP90. C’est bien connu, les protéines sont à la base de tous les mécanismes physiologiques, des nerfs aux muscles en passant par le sang, les hormones etc. Ce qui l’est moins, c’est que la fonction particulière de chaque protéine tient moins à sa composition chimique qu’à sa forme dans l’espace, à la manière compliquée dont elle se replie sur elle-même. Les protéines agissent en s’emboitant les unes dans les autres, à la manière d’un jeu de clés et de serrures. Mal configurée, une protéine est donc au mieux inactive, au pire source de dysfonctionnement. Il est donc vital que toutes les chaines protéiques soient parfaitement pliées sur elles-mêmes dès leur naissance. Or ces exercices de pliage n’ont rien d’évident car ils s’exécutent dans un milieu cellulaire totalement encombré par un tas de matériel organique. Un peu comme si on vous demandait de faire des pirouettes dans le RER à l’heure de pointe. Pas étonnant que près d’un tiers des protéines se loupe à un moment ou à un autre dans ces exercices de pliage. Heureusement, il existe dans les cellules des bonnes âmes pour aider les protéines en formation à bien plier leurs affaires, corriger leurs faux-plis et les empêcher de se lier à n’importe qui. Ces molécules “baby-sitters” dont HSP fait partie, “chaperonnent” en quelque sorte les bébés protéines, d’où leur nom de “protéines chaperons”. Elles les maintiennent aussi en stand-by jusqu'à ce que ce soit le moment pour elles d'entrer en action. De vrais nounous je vous dis! En temps normal, HSP90 et ses copines “canalisent” efficacement le développement des protéines en volant à leur secours dès que leur croissance est mise en péril par les fluctuations de l’environnement, une mutation inopportune ou les aléas du développement lui-même.
Un condensateur évolutif?
Un condensateur évolutif?
Si le stress est trop fort, sous l’effet d’un coup de chaud par exemple (HSP voulant dire “Heat Shock Protein”, protéine de choc thermique) ou si l’on inhibe chimiquement HSP90 (comme dans l’expérience en question), les erreurs de pliage s’accumulent et les super-nannies, débordées, laissent passer des protéines anormales et notamment celles qui reflètent les mutations masquées. Voilà pourquoi on obtient la même variabilité phénotypique en cas de stress thermique et d’inhibition chimique de HSP90! Pour Lindquist, HSP90 agirait donc comme un “condensateur évolutif” (“evolutionary capacitor”), permettant aux organismes de constituer des réservoirs de mutations “silencieuses”, bien utiles en cas de coup dur, à la manière d’un condensateur électrique accumulant les charges électriques et les libérant plus tard.

L’hypothèse est séduisante mais reste encore controversée car même si les expériences de Lindquist ont fait couler beaucoup d’encre, les validations expérimentales sont encore peu nombreuses. Certains chercheurs (voir cet article) ont par exemple émis l’hypothèse que la canalisation du développement serait une propriété inhérente à la complexité du système lui-même, sans qu’il y ait besoin d’un mécanisme canalisateur particulier. Autrement dit les très nombreuses interdépendances entre les gènes assureraient inévitablement une très grande robustesse du résultat final par rapport aux petites variations génétiques, un peu comme le réseau internet est à l’abri d’un gigantesque collapse du fait de son architecture distribuée. Selon cette métaphore, HSP90 ne serait pas la clé de voûte du système mais simplement un noeud vital du réseau, un peu comme le serait Google s'il était sur un seul site (thanks god ce n’est pas le cas!).
HSP90 contre les toxiques
L’autre critique porte sur le caractère adaptatif du mécanisme proposé par Susan Lindquist. Son expérience a été répétée avec succès sur les plantes et même les levures, suggérant que ces protéines chaperons sont un mécanisme évolutivement très très ancien (plus de 800 millions d’années d’évolution sépare la drosophile de la levure!).
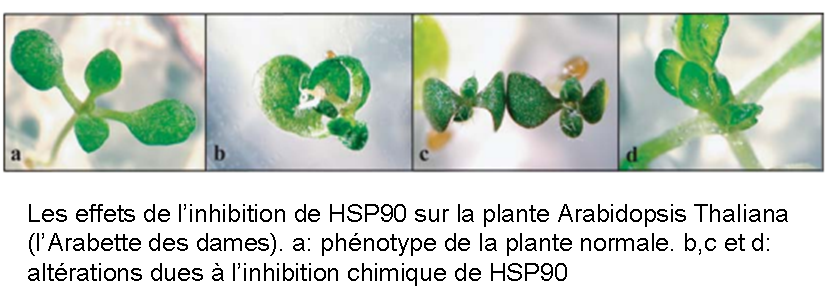 |
| (Source: Queitsch, Sangster et Lindquist, 2002, pdf) |
Or on voit mal comment l’évolution aurait pu sélectionner un caractère aussi abstrait que "la capacité à tamponner les mutations génétiques", dont la fitness se manifesterait au mieux une fois tous les mille ans. Peut-être s’agirait-il alors d’un effet collatéral de l’évolution, un “by-product”? Il se trouve justement que Lindquist (encore elle!) a découvert en 2005 un autre rôle-clé de HSP90 dans la capacité de résilience des cellules en cas d’agression externe. Les cellules cancéreuses par exemple, deviennent très rapidement résistantes aux médicaments employés en chimiothérapie, ce qui diminue l’efficacité de ces traitements. Pareil pour les cellules de levure, cultivées sur les mêmes substances toxiques. Mais -surprise!- Lindquist découvrit que la résistance des levures diminuait brutalement lorsqu’elle inhibait chimiquement HSP90 sur ces cellules.
 |
| (Source: Coweit et Lindquist 2005, pdf) |
Epigénétique ou pas?
Pour en revenir au rôle de HSP90 dans l’évolution, il reste quand même à savoir comment l’état mutant -révélé par le stress puis sélectionné pour sa fitness- se stabilise dans les générations successives, même en l’absence de stress. La question de l’hérédité épigénétique reste encore très largement ouverte. Pour Lindquist, ça n’a rien de très surprenant: les gènes ne constituent qu’une toute petite part de la matière organique de la première cellule embryonnaire. Le reste étant directement hérité de la mère, il ne serait pas complètement étonnant que certains caractères épigénétiques puissent se transmettre. Le biologiste Denis Noble remarque d’ailleurs que dans le monde pluricellulaire, l’hérédité des caractères acquis n’est pas l’exception mais la règle! Les cellules transmettent une grande partie de leur spécialisation à leur descendance, sans que leur bagage génétique soit modifié. Il n’y a guère que les cellules-souches totipotentes et les gamètes qui fassent exception.
Et puis, même si les effets de l'héritage épigénétique disparaissent au bout de quelques générations, peut-être suffisent-ils pour laisser le temps aux mutations classiques de prendre le relais et fixer génétiquement les caractères favorables selon les règles darwiniennes très classiques.
A défaut d’apporter toutes les réponses, Susan Lindquist a eu l'immense mérite d’ouvrir de nouvelles voies dans un tas de domaines de la biologie. Et encore je ne vous ai pas parlé du rôle des protéines chaperons dans le développement de certaines maladies mentales (Parkinson, Alzheimer...) ou des prions, toutes associées à des histoires de protéines mal repliées sur elles-mêmes. Décidément l’origami est une mine d'or pour la science!
Sources:
Le fameux article de Rutherford & Lindquist: HSP90 as a capacitor for morphological evolution (1998, pdf)
Cowen et Lindquit: Hsp90 Potentiates the Rapid Evolution of New Traits: Drug Resistance in Diverse Fungi (2005, pdf)
Un très bon exposé de la problématique par Vincent Debat, du MNHN (pdf)
Sur la résistance des levures: l’article de Science Daily: Heat shock protein drives yeast evolution (2010)