Les débats suscités par mon billet sur la "plasticité du vivant" m'ont donné envie de creuser le sujet et notamment d’explorer la piste évo-dévo suggérée par Taupo IRL. J'ai découvert que cette question de plasticité, loin d'être anecdotique, constitue un des sujets les plus chauds du moment en matière de biologie évolutive et pose un regard nouveau sur le rôle de la génétique. Récit en trois épisodes...
Repartons du schéma géno-centrique...
Aux yeux du grand public qui comme moi a lu (trop vite peut-être) le Gène Egoïste de Dawkins, la théorie synthétique de l’évolution a comme seul point de départ les mutations génétiques aléatoires, dont les effets sur les organismes sont soumis à la dure loi de la sélection naturelle. Dans ce schéma géno-centrique, l’évolution fonctionne à sens unique:
Repartons du schéma géno-centrique...
Aux yeux du grand public qui comme moi a lu (trop vite peut-être) le Gène Egoïste de Dawkins, la théorie synthétique de l’évolution a comme seul point de départ les mutations génétiques aléatoires, dont les effets sur les organismes sont soumis à la dure loi de la sélection naturelle. Dans ce schéma géno-centrique, l’évolution fonctionne à sens unique:

Comme l’explique Dawkins (p19)
les gènes réplicateurs “fourmillent maintenant en grandes colonies, à l’abri de gigantesques et pesants robots (les organismes, NDLA), isolés du monde extérieur, communiquant avec lui par des voies tortueuses et indirectes, et le manipulant par commande à distance.”Dans sa vision, les individus (les “robots”) sont de simples véhicules passifs et l’environnement n’a d’autre rôle que celui de filtrer en bout de chaîne les phénotypes les plus adaptés. En aucun cas il ne peut être directement la source d’innovations évolutives car ce privilège est réservé aux mutations génétiques aléatoires.
L’idée de “programme génétique” a marqué les esprits, dans une époque fascinée par les promesses de l’informatique naissante. C’est sans doute la raison pour laquelle elle continue de faire recette, malgré ses nombreuses limites. Car après tout, quel drôle de programme que celui de l’ADN, qui produit lui-même la machine capable de le lire! L’ADN n’est qu’une longue molécule qui dépend de toute la machinerie cellulaire pour que certains morceaux (les gènes justement) puissent être traduits en protéine. Et puis comme on va le voir, son code est loin d’être univoque: contrairement à un CD ROM qui n’a qu’une façon d’être interprété, il y mille et une façon de lire (ou pas) nos gènes...
Un génotype, plusieurs phénotypes
La métamorphose du stade larvaire au stade adulte montre bien qu'un même génotype peut se traduire en deux phénotypes différents. Un asticot ne ressemble pas franchement à une mouche quand même! Certes, on peut toujours imaginer que le programme génétique intègre plusieurs phases de développement qui sont toutes sous son contrôle. Mais comme ce “contrôle” se fait via des médiateurs chimiques (des hormones en général), il suffit que le milieu dans lequel baignent les cellules soit perturbé par l’extérieur pour que tout le développement soit chamboulé. C’est exactement ce qui se passe chez les axolotl, ces adorables pokemon d’eau qu’on trouve au Mexique et dont j’ai eu la chance de visiter un élevage à Xochimilco.
 |
| Un axolotl vu de mes yeux vu! |
“Les premiers axolotls importés en Europe au xixe siècle, furent installés au Jardin des Plantes de Paris. Ignorant leur néoténie, le professeur Auguste Duméril, le créateur du vivarium, fut un jour surpris de voir dans le bassin une espèce inconnue ressemblant à une salamandre à la place de l'animal qui s'y trouvait.”
 |
| Un axolotl après sa métamorphose |
 |
| Un tétard en flagrant délit de cannibalisme (source ici) |
Les exemples de polyphénisme (la capacité d’un même génotype à produire différents phénotypes) sont légions. Certains têtards de crapauds vivant dans les déserts américains sont capables de se métamorphoser en monstres cannibales lorsque le niveau d’eau baisse trop vite. Ils bouffent alors leurs frangins restés normaux pour faire le plein de calories avant de se métamorphoser en adultes. Une sorte de métamorphose de la métamorphose en quelque sorte.
J’aime bien aussi les transformations des criquets pèlerins: dès qu’ils se retrouvent en bandes, ils changent radicalement de forme et de comportement au point que l’on a longtemps cru qu’il s’agissait d’espèces différentes. Ils forment alors de gigantesques essaims volants qui peuvent ravager les cultures sur des centaines de kilomètres. Pour passer d'un inoffensif criquet solitaire à un criquet-garou, il suffit de lui chatouiller les pattes arrière!
De tels guilis simulent la proximité d’autres criquets qui le bousculeraient ce qui provoque une soudaine sécrétion de sérotonine dans certaines parties du corps et déclenchent sa métamorphose.

On pourrait continuer longtemps la liste des espèces qui se métamorphosent sous l’effet d’un changement de milieu:
- une alimentation particulière, pour les différentes castes de fourmis ou de termites- des variations de température, pour le sexe des alligators
- de l’humidité ambiante, pour la couleur et les motifs des ailes de papillons etc.
Dans la plupart des cas, on a réussi à reproduire artificiellement ces transformations, soit en contrôlant les conditions de l’environnement, soit en mettant directement les organismes au contact des bons médiateurs chimiques.

Alors pour qui votez-vous à ce concours de l’interaction entre organismes et environnement? Perso, ma préférence va au calmar bioluminescent (Euprymna scolopes) tel que le relate Jean-Claude Ameisen (“Dans la lumière et les ombres” p. 316):
“Un calmar nouveau-né possédera bientôt un organe lumineux qui lui permettra de chasser dans les ténèbres. Si cet organe émet de la lumière, c’est parce qu’il est habité par des bactéries marines luminescentes, des Vibrio fischeri”.
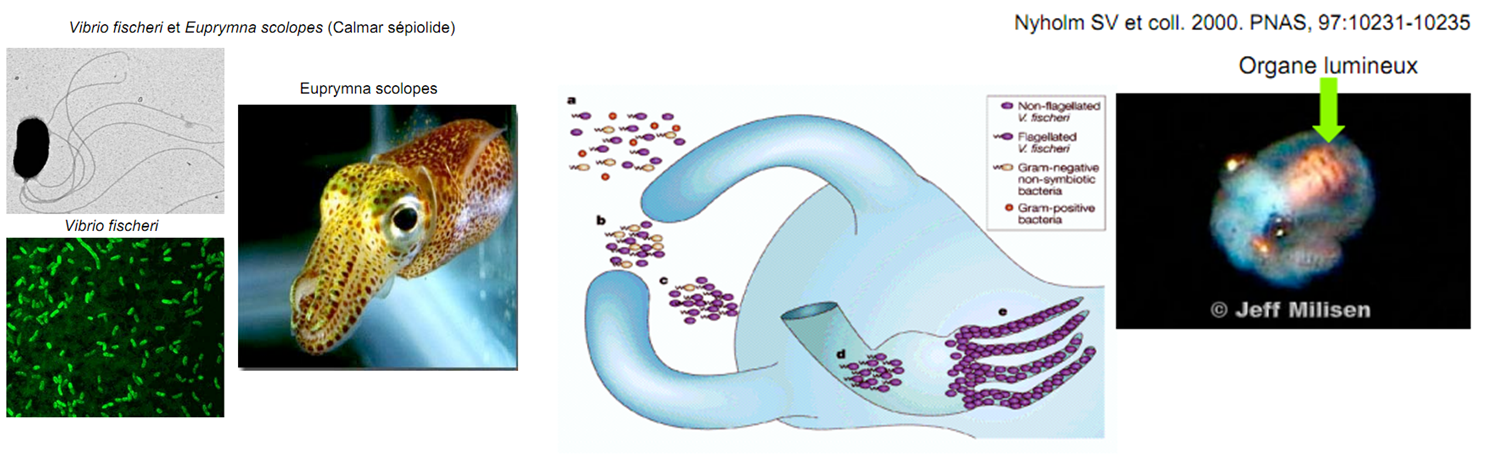
Chez le jeune calmar, l’organe bioluminescent ne se développe qu’à condition de recueillir des bactéries capables d’émettre de la lumière (sinon sa formation s’interrompt). En leur présence, les cellules qui tapissent la surface de l’organe s’autodétruisent pour laisser la place aux bactéries. De leur côté les bactéries ne peuvent émettre de la lumière que si elles se trouvent en concentration suffisante. La bioluminescence est donc aussi un phénomène collectif. Mais ce n’est pas tout! Une fois à l’abri de leur hôte, les bactéries perdent leur flagelle et deviennent ainsi un organe à part entière du calmar. A chaque génération, ce double mécanisme symbiotique se répète et avec lui cet incroyable dialogue entre deux organismes étrangers qui règlent leur physiologie l’un sur l’autre. Vous trouverez plein d’autres exemples de bioluminescence sur le blog de SSAFT, of course.
La réaction à l’imprévu
On pourrait objecter à tous ces exemples que les aléas en question sont suffisamment fréquents pour être pris en compte et mis à profit par un programme génétique optimisé après des millions d’années d’expérience. Il faut donc se tourner du côté d’aléas vraiment imprévisibles, donc hors de portée du mécanisme de sélection naturelle, pour savoir à quoi s’en tenir. Les anomalies de naissance sont un bon exemple de tels aléas.
 |
| Détails de l'anatomie de la chèvre à deux pattes - Cliquer pour aggrandir (source ici) |
Dans les années 1940, le cas d’une chèvre née par accident sans membre supérieur s’est avéré particulièrement intéressant. La pauvre bête fut bien obligée de se déplacer sur ses pattes arrière, comme un kangourou. Elle s’en sortait plutôt pas mal jusqu’à ce qu’elle meure accidentellement. L’analyse de son anatomie (qu’on peut lire chez PZ Myers) aurait fait le bonheur de D’Arcy Thompson, tant sa morphologie montrait une extraordinaire adaptation à sa condition de bipède: de plus grandes pattes postérieures (a), une colonne vertébrale incurvée et un cou anormalement long. L’anatomie (d) et la musculature pelvienne (b) étaient profondément modifiées du fait de la position verticale et l’on y retrouvait certaines des caractéristiques observées chez le kangourou... Pourtant cette chèvre était génétiquement tout à fait ordinaire. Les modifications de son phénotype étaient donc uniquemement induites par le stress physique provoqué par une posture verticale. C’est dire à quel point la plasticité phénotypique est grande et l’expression génétique imprévisible, tant elle dépend des interactions avec l’environnement. A défaut d’avoir une image de cette chèvre-kangourou, voici deux vidéos avec des cas semblables: celui d’une chèvre sans pattes arrières et d’un chien sans pattes avant.
 |
| La statue d'Alison Lapper artiste victime des effets de la Thalidomide, exposée sur Trafalgar Square (source ici) |
Bien sûr on pourra objecter que les adaptations morphologiques de cette pauvre bête à son handicap n’étaient pas transmissibles héréditairement et surtout qu’elles illustrent simplement à quel point la sélection naturelle a bien fait son boulot, avec des os, des tendons et des muscles conçus pour s’ajuster aux stress mécaniques qu’ils subissent. Je ne connais pas la raison pour laquelle la chèvre est née difforme, mais on sait bien que la prise de médicaments comme le Distilbène ou le Thalidomide pendant la grossesse peut provoquer de terribles malformations congénitales. Ces déformations observées parfois sur plusieurs gL'éénérations témoignent à l’évidence de la sensibilité de nos gènes (ou de leur expression) à l’environnement. Et elles nous rappellent au passage que cette influence n’est pas toujours adaptative (même si comme dans le cas de la chèvre à deux pattes, l’organisme compense très certainement une partie du handicap au cours du développement).
Les rythmes étranges de l’évolution
A l’autre bout de l’échelle, il est parfois possible de voir l’évolution à l’oeuvre. J’en ai donné un exemple avec les criquets Teleogryllus oceanicus, parasités par un insecte qui les repère au son de leur stridulation. En moins de vingt générations certains criquets ont adopté la morphologie des ailes de femelles, ce qui ne leur permettait plus de striduler. Pour compenser, ces petits malins se placent près des individus non mutants (qui eux continuent à striduler bêtement) et profitent comme ça des femelles sans se faire parasiter.
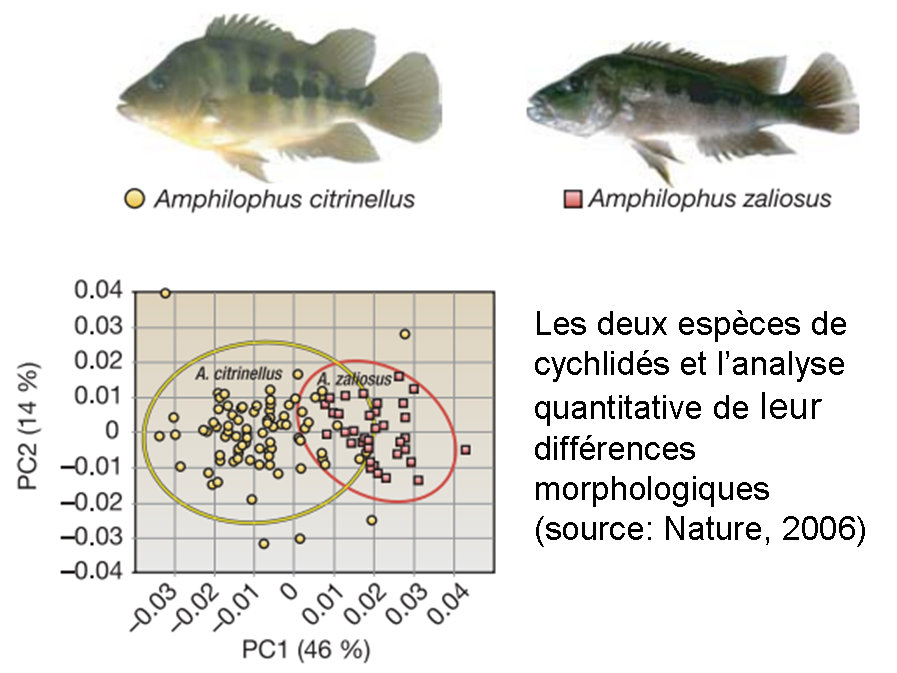
L’évolution ne se montre pas non plus très patiente avec les cychlidés, ces magnifiques poissons tropicaux qui font le bonheur des aquariophiles et qui semblent capables de se diversifier comme qui rigole. Cet article de Nature raconte comment l’espèce endémique (Amphilophus citrinellus) introduite il y a 23 000 ans dans un lac nouvellement formé du Nicaragua a mis moins de 10 000 ans pour diverger et se transformer en une tout autre espèce (Amphilophus zaliosus), alors qu’elle restait immuable dans les lacs des alentours.
L’évolution ne se montre pas non plus très patiente avec les cychlidés, ces magnifiques poissons tropicaux qui font le bonheur des aquariophiles et qui semblent capables de se diversifier comme qui rigole. Cet article de Nature raconte comment l’espèce endémique (Amphilophus citrinellus) introduite il y a 23 000 ans dans un lac nouvellement formé du Nicaragua a mis moins de 10 000 ans pour diverger et se transformer en une tout autre espèce (Amphilophus zaliosus), alors qu’elle restait immuable dans les lacs des alentours.
 |
Ces exemples apportent deux enseignements:
- Ils montrent d’abord que le mécanisme naïf envisageant l’évolution comme l’attente passive d’une mutation salvatrice ne tient pas toujours la route. Le mécanisme de l’évolution, le fameux ’”horloger de Dawkins” est peut-être aveugle, mais il n’est manifestement pas complètement sourd puisqu’il sait accélérer ses essais hasardeux lorsqu’on le stresse un peu. On peut donc imaginer qu’un système de feedback entre l’environnement et le taux de mutations exprimées permet aux organismes de s’adapter plus vite en cas de besoin.
- Ensuite, le contraste est frappant entre la versatilité des espèces citées plus haut et l’immuabilité génétique et morphologique d’autres comme le Ginkgo (240 millions d’années) ou le cœlacanthe (350 millions d’années. Respect!).
- Ils montrent d’abord que le mécanisme naïf envisageant l’évolution comme l’attente passive d’une mutation salvatrice ne tient pas toujours la route. Le mécanisme de l’évolution, le fameux ’”horloger de Dawkins” est peut-être aveugle, mais il n’est manifestement pas complètement sourd puisqu’il sait accélérer ses essais hasardeux lorsqu’on le stresse un peu. On peut donc imaginer qu’un système de feedback entre l’environnement et le taux de mutations exprimées permet aux organismes de s’adapter plus vite en cas de besoin.
- Ensuite, le contraste est frappant entre la versatilité des espèces citées plus haut et l’immuabilité génétique et morphologique d’autres comme le Ginkgo (240 millions d’années) ou le cœlacanthe (350 millions d’années. Respect!).
 |
| L'arbre Ginkgo (source ici) |
 |
| Coelacanthe (source ici) |
Le gène propose, l’individu dispose...
Voici ce que pourrait donner le système de l’évolution révisé:

1) L’environnement est maintenant au centre du schéma et influence à peu près tous les autres niveaux. Entre acquis et inné, la différence devient donc ténue car on pourrait presque dire que le gène propose et l’environnement dispose...
2) Le lien entre génotype et phénotype passe manifestement très souvent par une étape critique, celle du développement (de préférence durant la phase embryonnaire mais parfois aussi chez l’adulte) durant laquelle se joue un enchevêtrement hyper complexe d’interactions entre tous les niveaux et dont émerge l’incroyable plasticité du phénotype. On est à l’opposé de ce qu’affirmait Dawkins pour qui “les détails du développement embryonnaire environnemental, aussi intéressants qu’ils soient, sont inutiles du point de vue de l’évolution” (p62).
3) Dans ce schéma, le pouvoir des gènes est considérablement atténué. Comme le remarque malicieusement Didier Noble (La musique de la vie, p34), on pourrait remplacer la sentence de Dawkins par son contraire, ce serait tout aussi arbitraire:
“Les gènes sont aujourd’hui piégés en grandes colonies, enfermés dans des êtres hautement intelligents, façonnés par le monde extérieur, comuniquant avec lui par des processus complexes par elsquels, aveuglément, comme par magie, la fonction émerge. Ils sont en vous et en moi; nous sommes le système qui permet à leur code d’être lu”.Bon, reste à comprendre quels sont les mécanismes moléculaires qui permettent cette plasticité du phénotype et des mutations. Ce sera pour le prochain billet!
Sources:
Dawkins, Le gène égoïste (1990)
Denis Noble, La musique de la vie (2010)
… et tous les billets cités dans le billet.
Billets connexes:
Plastique la vie! L’objet des débats...
Innovations, scrabble et logarithmes: l’évolution, une histoire de combinatoire sous contrainte?
Innovation et évolution: une histoire de lapins sauteurs? sur les étranges ramifications de l’arbre de la vie
L’évolution c’est de la dynamique, sur le rôle moteur des contraintes biomécaniques au cours du développement.